 치과건강보험
치과건강보험
 치과경영칼럼
치과경영칼럼
 법률칼럼
법률칼럼
 직원관리
직원관리
 마케팅
마케팅
Ⅱ. 아편의 효과와 수용체 및 길항제 (4)

6. 도파민과 도파민 수용체
도파민은 영국, 런던의 Wellcome 연구소에서‘George Barger’와‘James Ewens’에 의해 1910 년에 종합되었다. 기본적 구조가 모노아민이기 때문에 도파민이라 불리게 되었다. 전구물질은‘Barger-Ewens’합성에서 3, 4-dihydroxyphenylalanine(levodopamine 또는 L- DOPA) 이다.
인간의 경우, 도파민은 음식, 섹스, 마약과 같은 일부 약물과 자극으로 보상의 경험에 의해 분비되는 물질이지만 흡연으로 인해 흡수되는 니코틴도 모르핀 효과와 유사하게 도파민의 유 리를 활성화 시켜서 쾌감을 느끼게 해준다. 1910년에 영국 런던의 Wellcome 연구소에서 ‘George Barger’ 와‘James Ewens’ 에 의해 도파민이라는 새로운 신경 전달물질의 존재가 발표되었다. 그것은 일종의 모노아민 화합물이었기 때문에 도파민이라 불리게 되었다.
도파민의 전구물질은 Barger - Ewens 합성에서 3, 4-dihydroxyphenylalanine (levodopamine 또는 L-DOPA) 이다. 신경전달물질로서의 도파민의 기능은 처음 1958년 ‘Arvid Carlsson’ 과 ‘Nils-Åke Hillarp’ 에 의해 스웨덴 국립심장연구소화학, 약리학 연구파트에서 발견되었다. ‘Carlsson’은 노아에피네프린과 에피네프린의 전구물질일 뿐 아니라 신경전달물질로서의 도파민의 기능을 밝혀냄으로써 2000년에 노벨상을 수상했다.
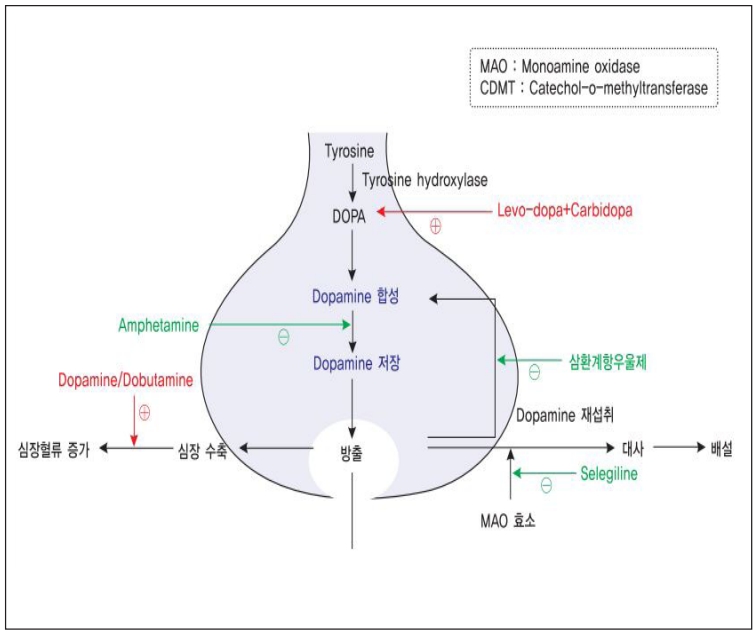
도파민의 생성 및 분해과정에는 두 가지의 주요 경로가 존재한다. 선조체와 대뇌기저핵을 포함한 뇌의 대부분의 구역에서 도파민은 도파민전달(DAT1)후 두 가지의 모노아민 옥시다제 (MAOA와 MAOB)에 의해 3, 4-dihydroxyphenylacetic 산으로 효소분해를 통해 불활성화된다. 그렇지만 뇌전두엽피질에는 도파민 전달 단백질이 거의 없기 때문에 도파민은 대신 norepinephrine 전달(NET)후 인근 norepinephrine의 뉴런에서 효소분해 (COMT)를 통해 3-methoxytyramine로 불활성화된다.
DAT1 경로는 NET 경로보다 빠르다. 마우스에서 도파민 농축은 꼬리가 있는 핵에서는 200ms의 반감기 (DAT1 경로를 사용하는) 대비 전두엽피질에서는 2,000ms로 사라진다. 효소에 의해 분해되지 않는 도파민은 VMAT2에 의해 재사용을 위해 vesicles로 재합성된다. ‘Ming-Hu Han’ 과‘Karl Deisseroth’ 의 실험실에서 발표한 두 편의 연구논문에서는 스트레스에 의한행동에 대해 도파민이 신경세포를 조절하는 효과에 대한 내용이 발표되었다.
‘Han’ 과 공동연구원들은 급격한 스트레스 경험에 의한 사회성 결여 시 나타나는 도파민의 활성패턴 차이의 기능적 효과에 대한 연구결과를 발표하였던 것이다. ‘Deisseroth’ 와 공동 연구원들은 장기간의 스트레스 상황에 노출된 경우에 유발되는 행동학적 효과 및 신경회로 변경에 대한 VTA 뉴런 활성을 조절하는 도파민의 효과에 대해서 연구하였다.
인용문헌: ‘치과처방의 완성’(대한민국학술원 선정, 교육부 지정 2020 우수학술도서)

김영진 박사
건강보험심사평가원 상근 심사위원 역임
보건복지부장관 위촉 금연진료의료인 교육강사